
Структура политенных хромосом и морфология личинок хирономид (Diptera, Chironomidae). Атлас
Покупка
Новинка
Тематика:
Микробиология
Издательство:
КМК
Год издания: 2022
Кол-во страниц: 271
Дополнительно
Вид издания:
Монография
Уровень образования:
ВО - Магистратура
ISBN: 978-5-907533-61-5
Артикул: 823156.01.99
Доступ онлайн
439 ₽
В корзину
Монография посвящена анализу кариотипов 109 видов хирономид (Chironomidae, Diptera) Палеарктики и некоторых других регионов. Приводятся данные, полученные авторами в течение десятилетий работы в области изучения хромосом хирономид. Изученные виды относятся к подсемействам Tanypodinae (5 родов, 6 видов), Diamesinae (3 рода, 11 видов), Prodiamesinae (1 род, 2 вида), Orthocladiinae (4 рода,
12 видов) и Chironominae (18 родов, 78 видов). Кариотипы 9 видов описываются впервые. Для большинства видов приводятся оригинальные цитофотокарты политенных хромосом и морфологические описания личинок.
Монография рассчитана на цитогенетиков, кариологов, гидробиологов, энтомологов, экологов. Она также может быть полезна преподавателям и студентам биологических вузов.
Тематика:
ББК:
УДК:
ОКСО:
- ВО - Бакалавриат
- 00.03.12: Экология
- 06.03.01: Биология
- ВО - Магистратура
- 06.04.01: Биология
- ВО - Специалитет
- 00.05.12: Экология
ГРНТИ:
Скопировать запись
Фрагмент текстового слоя документа размещен для индексирующих роботов.
Для полноценной работы с документом, пожалуйста, перейдите в
ридер.
�����������������������������������������������������������������������������������������������������������������������������������������������������������������������
Светлой памяти Л.А. Чубаревой (1921–2006) Предисловие Хирономиды, или комары-звонцы (Chironomidae) — одно из широко распространённых семейств двукрылых насекомых (Diptera). Его представители обитают во всех зоогеографических областях, включая Антарктическую (Martin, 1962; Atchley, Davis, 1979; Butler, 1982; Белянина, Филинкова, 1996, 2014; Крашенинников, 2013). Хирономиды встречаются практически во всех типах водоёмов, а также в почве, навозе, грибах, дуплах деревьев, гниющих корягах и влажном мху. Значительное число видов живет в прудах, озерах, заливах и литоральных участках морей, в солёной и солоноватой среде (Шоба- нов, Дёмин, 1988; Orel (Zorina) et al., 2014; Маркиянова, 2017; Маркиянова, Петрова, 2020). В пресноводной фауне насекомых нет другой группы, которая была бы представлена таким разнообразием видов, характеризующихся столь же высокой численностью и экологической пластичностью, как хирономиды. Высокая экологическая пластичность проявляется, например, в том, что личинки видов, обитающих в Африке, с наступлением сухого сезона формируют защитный кокон для предотвращения значительной потери воды и впадают в диапаузу (МсLachlan, 1983а, b), находясь в высохшей грязи в аметаболическом состоянии (ангидробиоз) до наступления сезона дождей. В таком состоянии личинки сохраняются при температурах от –27 °С до +120 °С и могут активироваться, попав в воду, даже через 18 лет (Hinton, 1951, 1960а, b; Adams, 1985). Хозяйственное значение хирономид очень велико. Личинки являются основным, а иногда и единственным кормом для промысловых бентосоядных рыб, хищных насекомых и птиц (Шилова, 1976). Иногда количество хирономид в водоеме столь значительно, что желудки рыб заполнены только ими. Хирономи- ды участвуют в самоочищении водоемов, перерабатывая органические вещества в минеральные. Личинок хирономид также используют в качестве индикаторов степени загрязнения рек и трофности озер (Макрушин, 1974; Тодераш, 1984; Балушкина, 1987), в частности, индикаторов степени загрязнения водоемов радионуклидами, тяжёлыми металлами и др. (Петрова, 2013). Среди хиро- номид известны эктопаразиты насекомых, моллюсков и рыб. Иногда имаго хи- рономид вызывают у людей различные формы аллергии (бронхиальную астму, сезонные риниты и конъюнктивиты), а на юге России личинки некоторых видов зарегистрированы как вредители риса (Шилова, 1986).
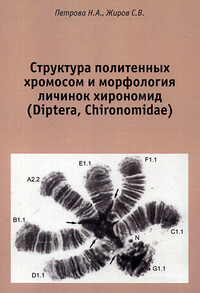
Н.А. Петрова, С.В. Жиров Структура политенных хромосом… В семействе существует до сих пор нерешённая проблема — несоответствие количества видов, описанных по личинке или куколке, числу видов, описанных по имаго. Это объясняется, во-первых, тем, что первые исследователи описывали виды, основываясь зачастую на сухом коллекционном материале, преимущественно по окраске, не используя при этом некоторые структурные признаки, имеющие таксономическое значение. Во-вторых, энтомологи описывали (и до сих пор описывают) виды по имаго, а гидробиологи — преимущественно по преимагинальным водным стадиям; при этом классификация по личинкам и куколкам большей частью не совпадает с классификацией по имаго (Thienemann, 1922; Edwards, 1926; Goetghebuer, 1928). В-третьих, личинки нескольких видов могут относиться к одному и тому же морфологическому типу. В большинстве случаев определение личинок из-за их морфологического единообразия (недостаточного количества значимых таксономических признаков) оказывается возможным только до группы видов. Мы столкнулись с подобной ситуацией, например, в роде Cricotopus van der Wulp, 1874, в котором по кариотипическим признакам выделяются два дискретных варианта, соответствующих, предположительно, одному виду (Petrova et al., 2012). Наконец, в семействе существует, по-видимому, много близких видов, которые можно надёжно различать только по кариотипу (например, Glyptotendipes paripes Edwards, 1929, Chironomus pilicornis (Fabricius, 1794), Ch. frequentatus Belyanina et Filinkova, 1996 и др.). Последние 30 лет систематики-хирономидологи пытаются исправить положение и построить единую систему, которая бы учитывала не только морфологию личинки, но и структуру политенных хромосом. Эти хромосомы более чем в 200 раз превышают по размеру обычные митотические хромосомы (поэтому их ещё иногда называют гигантскими политенными хромосомами) и характеризуются чередованием поперечных темных и светлых полос, так называемым рисунком дисков, который видоспецифичен. Кариологический метод обладает большой разрешающей способностью и является надежным методом видовой идентификации личинок. С середины XX века и до настоящего времени он успешно применяется при решении трудных вопросов систематики и филогении хирономид (Keyl, Keyl, 1959; Keyl, 1962; Martin, 1979; Белянина, 1983; Michailova, 1989; Кикнадзе и др., 1991а, 1996a; Kiknadze et al., 2012, 2016; Петрова, 2013; Kaрмоков и др., 2014). Уникальное строение политенных хромосом позволяет идентифицировать не только каждую хромосому и небольшие её участки, но и любые структурные изменения кариотипа. Таким образом, политенные хромосомы с их видо- специфическим рисунком дисков служат базой для кариосистематического изучения семейства Chironomidae.
Условные обозначения Морфологические признаки: Кариологические признаки: Сокращения: AF — подставка антенны АR — индекс антенны (отношение длин базального членика и жгута) AS — анальная кисточка Bl — сенсилла антенны DZ — верхние зубцы мандибулы E — эпифаринкс LO — лаутерборнов орган LZ — боковые зубцы мандибулы M — ментум Mand — мандибула Max — максилла MP — максиллярный щупик MZ — срединный зубец мандибулы MZ1— основной зубец мандибулы Pc — подставка анальной кисточки PE — эпифарингеальный гребень Pm — премандибула PP — задние подталкиватели RO — кольцевой орган Sm — субментум SSd — щетинка под зубцами мандибулы TLt — латеральные отростки BR — кольцо Бальбиани P — пуфф Chr — хромоцентр Inv — инверсия B-хр. — В–хромосомы 2n — диплоидное число хромосом I, II и т.д. — номера хромосом A, B и т.д. — плечи хромосом 1, 2 и т.д. — районы хромосом N — ядрышко оз. — озеро окр. — окрестности о. — остров пос. — поселок р-н — район р. — река руч. — ручей сев. — северный с. — село ст. — станция водохр. — водохранилище возвыш. — возвышенность вост. — восточный гл. — глубина дер. — деревня зал. — залив зап. — западный м. — море над у.м. — над уровнем моря обл. — область
Благодарности Авторы выражают искреннюю признательность научным редакторам профессору, д.б.н. В.Г. Кузнецовой и к.б.н. Н.В. Голуб, рецензентам д.б.н. В.Ф. Шуйскому и к.б.н. С.А. Галкиной, д.б.н. С.Ю. Синёву, д.б.н. Б.А. Коро- тяеву и д.б.н. E.В. Даеву за ценные советы по оформлению рукописи, а также к.б.н. А.А. Пржиборо за критические замечания. Авторы благодарны Л.А. Чу- баревой, В.Я. Панкратовой, П. Михаиловой (P. Michailova, Болгария) и сотрудникам отделения кариосистематики лаборатории систематики насекомых Зоологического института РАН и, прежде всего, руководителю отделения В.Г. Кузнецовой за поддержку работы, научные дискуссии и дружескую атмосферу, а также Н.С. Хабазовой за помощь при оформлении рукописи, особенно её иллюстративной части. Авторы признательны всем отечественным и зарубежным коллегам, которые собирали материал, помогали его определять и были соавторами публикаций — М.В. и К.В. Арутюновым, Е.В. Балушкиной, С.И. Беляниной, Н.М. Бухтеевой, Е.Б. Виноградовой, Н.В. Винокуровой, О.А. Гусеву, М.В. Даниловой, Э.А. Ербаевой, О.В. Ермоловой, Н.И. Зеленцову, Н.Б. Ильинской, А.Г. Истоминой, М.Х. Кармокову, Э.А. Качворян, И.Е. Керкис, И.И. Кик- надзе, O.K. Клишко, А.Б. Крашенинникову, Н.К. Кузьменко, Л.А. Куприяновой, Л.Л. Левитиной, Л.Е. Лобковой, Е.А. Макарченко, Ф.Л. Максимовой, О.В. Орел, А.Ж. Ракишевой, Л.В. Фегер, Н.А. Шобанову, П. Адлеру (P. Adler, США), С. Боверо (S. Bovero, Италия), К. Брюнетти (K. Brunetti, Италия), М. Джинепро (M. Ginepro, Италия), Ю. Илковой (J. Ilkova, Болгария), О. Кави- чиоли (O. Cavicchioli, Италия), Т. Кикавада (T. Kikawada, Япония), Т. Кобаяши (T. Kobayashi, Япония), Р. Корнетту (R. Cornette, Япония), И. Матена (J. Matena, Чехия), Т. Окуда (T. Okuda, Малави), Д. Пемба (D. Pemba, Малави), Л. Раме- ла (L. Ramela, Италия), Г. Селла ( G. Sella, Италия), Д. Тодоровой (D. Todorova, Болгария), К. Уайту (K. White, Великобритания), Т. Ханкелну (T. Hankeln, Германия), С. Шимуре (S. Shimura, Япония), Е. Шмидту (E. Shmidt, Германия), Н. Ямамото (N. Yamamoto, Япония) и М. Ямамото (M. Yamamoto, Япония),
Ãëàâà 1 Внешнее строение личинки В главе приводится общая характеристика личинки (рис. 1) и указываются наиболее существенные для систематики морфологические признаки. Здесь (а также в Главе 2) использованы описания и термины, заимствованные из работ Панкратовой (1970, 1977, 1983), Макарченко (1980, 1985, 1998, 1999), Фиткау и Робак (Fittkau, Roback, 1983), Оливер (Oliver, 1983), Крэнстон и др. (Cranston et al., 1983), Зетер (Saether, 1980, 1983), Пиндер и Рейс (Pinder & Reiss, 1983). Важно, что, используя одну и ту же личинку, можно изучить ее внешнюю морфологию и кариотип. Особенно это целесообразно при работе с новыми и редкими видами (Макарченко, 1980; Saether, 2000). Личиночная стадия хирономид — самая длительная часть их жизненного цикла, в то время как куколки и имаго живут от нескольких недель до нескольких дней. Тело личинки червеобразное, состоит из хорошо оформленной головы, 3 грудных и 10 брюшных сегментов (рис. 1, а–в). На последнем брюшном сегменте имеются парные цилиндрические выросты тела — подталкива- тели, снабженные крючками (PP). На предпоследнем сегменте со спинной стороны расположены так называемые подставки преанальных кисточек, которые состоят из небольшого числа длинных жестких щетинок (Pc). На VII брюшном сегменте у некоторых видов Chironomus и Camptoсhironomus развиты латеральные отростки (TLt), на VIII — одна или две пары вентральных отростков (TV) разных размера и формы (Сhironomus, Camptoсhironomus, Glyptotendipes). Голова имеет наибольшее число видовых диагностических признаков (рис. 1, г–д). Одна из особенностей головы − наличие втяжных антенн, которые состоят обычно из 5–7 члеников (рис. 2, а–д), 1-й членик принято называть основным, или базальным, остальные все вместе составляют жгут, который претерпевает вторичное расчленение. На поверхности основного членика антенны имеются один или несколько кольцевых органов (рис. 2, а–в), а на его конце, рядом со жгутом, расположена расщепленная или простая сенсилла, которая может достигать вершины антенны. Важное значение имеет индекс антенны: отношение длины 1-го членика к длине всего жгута. Соотношение длин всех члеников антенны также видоспецифично. Обычно жгут антенны имеет 4, 5 или 6–7 члеников. На дистальном конце жгута имеются два противостоящих либо последовательных лаутерборновых органа, но эти органы могут быть и непарными. Ротовая полость располагается на нижней стороне головы, а сверху и спереди ограничена верхней губой (лабрум), на которой располагается эпифаринге- альный гребень (рис. 2, е). По широкому верхнему краю эпифаринкса расположены гребешки, состоящие из 3 частей, расположенных в один ряд (они могут быть одного размера, или крупные зубчики чередуются с мелкими), но у некоторых видов имеются многорядные зубчики. Строение эпифаринкса у большинства родов довольно однотипно (рис. 2, з). Верхние челюсти или мандибу-
Н.А. Петрова, С.В. Жиров Структура политенных хромосом… лы (рис. 3, а–б) соприкасаются с премандибулами (рис. 3, в–г). Мандибулы более или менее серповидно изогнуты и несут на внутренней поверхности, близ вершины, зубцы: нижние (три–четыре) обычно темные, хорошо развитые, и верхние (один–два) небольшие. Под зубцами с внешней нижней стороны мандибулы есть плоская щетинка, известная как «щетинка под зубцами» (рис. 3, б). На рис. 2, ж показана мандибула, встречающаяся у личинок примитивных таксонов. Максиллы представляют собой плоские лопасти, неподвижно причлененные к щёчным склеритам, и несут многочисленные щетинки (рис. 3, д), позади которых находится нижняя губа, или ментум (рис. 4, а–в). Передний край ментума зубчатый (рис. 4, а), причем форма, цвет, число и размер зубцов у разных видов различны. Их может быть четное или нечетное число; в первом случае принято различать два срединных зубца, во втором — один. Как правило, боковые зубцы постепенно уменьшаются от середины ментума к его бокам. Вентроментальные пластинки (рис. 4, б–в) чаще не соприкасаются друг с другом, но бывает и наоборот. Морфологические препараты личинок приготавливались по методикам отечественных и зарубежных специалистов (Черновский, 1949; Панкратова, 1970, 1977, 1983; Шилова, 1976; Макарченко, 1980, 1985, 1998, 1999; Fittkau, Roback, 1983; Oliver, 1983; Cranston et al., 1983; Pinder, Reiss, 1983).
Доступ онлайн
439 ₽
В корзину